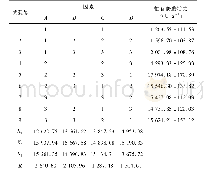
《表2 黑曲霉分泌酶:黑曲霉作为分泌蛋白细胞工厂的研究进展》
 提示:宽带有限、当前游客访问压缩模式
提示:宽带有限、当前游客访问压缩模式
(1)D、E、P、S和A分别表示通过直接分离纯化、大肠杆菌表达、毕赤酵母表达、酿酒酵母表达和黑曲霉表达获得目标蛋白。
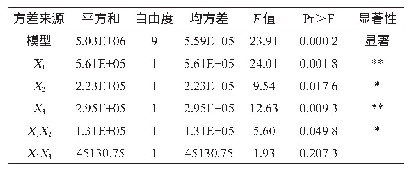
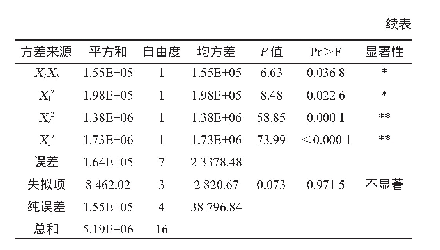
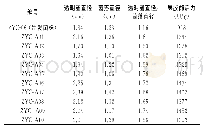
黑曲霉自身就是一个酶的宝库。黑曲霉分泌蛋白中包含了大量能够降解多糖、蛋白和脂肪的酶[30]。黑曲霉来源的纤维素酶、半纤维素酶、果胶酶、淀粉酶、菊粉酶、脂肪酶和蛋白酶等广泛应用于食品、饲料、洗涤剂、生物燃料等领域。目前,大量关于黑曲霉分泌酶性质和结构的研究被相继报道,这对酶的高效利用具有重要意义。表2总结了近十年来报道的部分黑曲霉分泌酶。可以看到,黑曲霉来源的纤维素酶和半纤维素酶占了很大的比例,这得益于木质纤维素降解酶在生物燃料中的应用。值得注意的是,表2中大部分酶都是通过从发酵液中直接分离纯化或者通过毕赤酵母异源表达得到的。直接分离纯化得到纯酶的方法适用于组分较为简单的发酵液。考虑到黑曲霉大部分分泌酶为诱导酶[31-33],并且在一个诱导条件下往往会产生一系列的酶,因此,要想直接分离得到纯酶,需要结合多步分离手段,包括硫酸铵沉淀、超滤、离子交换层析、疏水作用层析、体积排阻层析、聚焦电泳等,操作复杂,耗时耗力。此外,通过分析黑曲霉基因组范围的蛋白发现,在黑曲霉中涉及到木质纤维素降解的分泌酶就有131个[30],全部的分泌酶将近有900个[34-35]。通过二维电泳及质谱分析能快速获得蛋白种类信息,然而,通过实验得到的分泌酶种类往往会比理论分析得到的数据小一个数量级[34]。因此,在实际发酵中,黑曲霉分泌酶往往只是其基因组范围内分泌酶的小部分,大部分潜在的分泌酶未能在特定条件下表达,直接分离纯化的方法往往会错失很多具有优良特性的酶。过表达目的蛋白的方法很好地解决了上述问题。毕赤酵母作为真核表达宿主,具有较完备的蛋白折叠和修饰系统以及较强的蛋白分泌能力,能够胜任大部分真核分泌酶的异源表达[36]。当然,异源表达也存在弊端,即真核表达系统中蛋白糖基化修饰的差异[37]。糖基化修饰的差异对酶的性质有一定的影响,这不利于后续的研究和应用。此外,毕赤酵母在表达较大分子量的蛋白时效率往往很低,低的表达量会间接地增加后续分离纯化的难度。相比较而言,同源表达往往是最佳的选择。
| 图表编号 | XD00101432100 严禁用于非法目的 |
|---|---|
| 绘制时间 | 2019.10.01 |
| 作者 | 姚善泾、蔡礼年、林东强 |
| 绘制单位 | 浙江大学教育部生物质化工重点实验室化学工程与生物工程学院、浙江大学教育部生物质化工重点实验室化学工程与生物工程学院、浙江大学教育部生物质化工重点实验室化学工程与生物工程学院 |
| 更多格式 | 高清、无水印(增值服务) |

{kind=link}