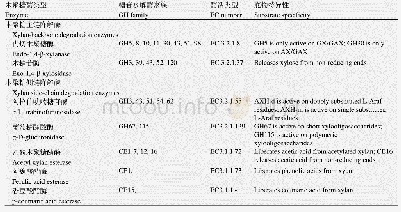
《表3 降解或淬灭QS的AHLs酰基转移酶》
 提示:宽带有限、当前游客访问压缩模式
提示:宽带有限、当前游客访问压缩模式
本系列图表出处文件名:随高清版一同展现
《基于AHLs介导的革兰氏阴性菌群体感应调控及淬灭机制研究进展》
在芽孢杆菌属240B1中发现AiiA内酯酶后不久,又发现菌株争论贪噬菌VAI-C可以AHLs作为能源和氮源诱导形成一种AHLs酰基转移酶,该酶不可逆地水解AHLs酰胺键后形成HSL和相应的脂肪酸(图5b)。争论贪噬菌VAI-C可以降解不同长度酰基侧链(从C4到C12)的各种饱和AHLs和3-oxo-C8-HSL。脂肪酸可作为碳源和能源支持VAI-C的生长,而HSL只作为氮源。后续研究在铜绿假单胞菌、青枯菌[61]、丛毛单胞菌属[67]、丁香假单胞菌[65]、希瓦氏菌属[60]、苍白杆菌属[64]、红串红球菌W2[39]和链霉菌M664[62](表3)等细菌中也鉴定了AHLs酰基转移酶[70]。布鲁氏菌是一种能够产生长链AHLs的导致人畜共患病的病原体,在体外或巨噬细胞感染期间可以产生具有控制内在AHLs自诱导剂的AHLs酰基转移酶(AibP),降解AHLs后影响其致病作用[63]。在铜绿假单胞菌中发现和表征了另一种AHLs酰基转移酶PvdQ,它与AiiD酰基转移酶具有很高的同源性,只降解长链AHLs。
| 图表编号 | XD00159705800 严禁用于非法目的 |
|---|---|
| 绘制时间 | 2020.08.31 |
| 作者 | 崔天琦、白凤翎、励建荣 |
| 绘制单位 | 渤海大学食品科学与工程学院生鲜农产品贮藏加工及安全控制技术国家地方联合工程研究中心、渤海大学食品科学与工程学院生鲜农产品贮藏加工及安全控制技术国家地方联合工程研究中心、渤海大学食品科学与工程学院生鲜农产品贮藏加工及安全控制技术国家地方联合工程研究中心 |
| 更多格式 | 高清、无水印(增值服务) |



{kind=link}