《表2 Q426菌株组氨酸激酶的结构域功能》
 提示:宽带有限、当前游客访问压缩模式
提示:宽带有限、当前游客访问压缩模式
本系列图表出处文件名:随高清版一同展现
《解淀粉芽孢杆菌Q426双组分信号转导系统的生物信息学分析》
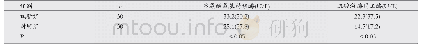
大部分的HKs是有个多结构域且能够形成同型二聚体的跨膜蛋白[12],HK的N-端存在多个跨膜区,不同结构域分布其中,使HK的结构和功能具有多样性;HK的C-端区域有多个ATP结合位点,典型的HK有HATPase_c和His KA结构域,HAT-Pase_c结构域位于胞内,包含4个保守的N、D、F和G-Box,是组氨酸激酶的催化结构域,主要负责把ATP的磷酸基团转移到His KA结构域的组氨酸残基上。His KA是磷酸受体结构域,该区域含有一个保守组氨酸位点的H-box,能够自身磷酸化[13]。从Pfam数据库中得知,目前已发现的结构域超过19 000种,因为结构域是一种介于二级和三级结构之间的一种结构层次,所以不同结构域组合的蛋白质表达不同的功能,比如:DegS结构域与细胞生长周期的调控有关,含有该结构域的HK可能参与调控细胞生长周期。在Q426菌株中找出的42个HKs含HAMP、PAS、PAC、s Cache_like、7TMR-DISM_7TM、REC、Cache_3、SPOB_a、HSP90、GAF、HPT、MutL_C、PDZ等25种结构域,其中保守性高的HisKA和HATPase_c是所有HKs必需的结构域,它们是磷酸基团传递的重要基础。这25种结构域的功能如表2所示。其中,19个成对的HK包含11种结构域,这19个HK的结构域组成如图1所示(图中左列数字是HK在基因组数据中的编号,右侧是HK的结构域组合)。
| 图表编号 | XD0075571000 严禁用于非法目的 |
|---|---|
| 绘制时间 | 2019.08.20 |
| 作者 | 师舷、刘佳璐、董极靓、吴亚多、权春善 |
| 绘制单位 | 大连民族大学生命科学学院生物技术与资源利用教育部重点实验室、大连民族大学生命科学学院生物技术与资源利用教育部重点实验室、大连民族大学生命科学学院生物技术与资源利用教育部重点实验室、大连民族大学生命科学学院生物技术与资源利用教育部重点实验室、大连民族大学生命科学学院生物技术与资源利用教育部重点实验室 |
| 更多格式 | 高清、无水印(增值服务) |





{kind=link}