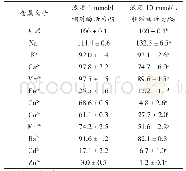
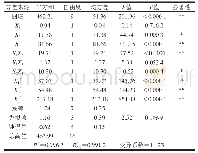
《表1 多糖降解酶活性改造的实例》
 提示:宽带有限、当前游客访问压缩模式
提示:宽带有限、当前游客访问压缩模式
1)在最适条件下测定的各酶分子突变前后的比酶活.2)在最适条件下测定的各酶分子突变前后的催化效率.
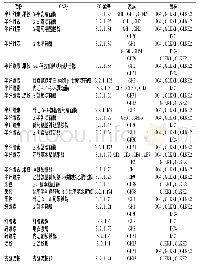
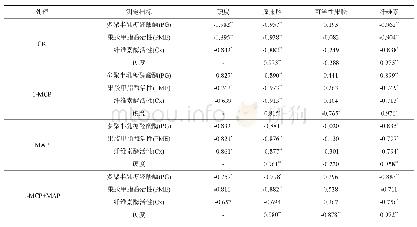
碳水化合物结合模块(CBM)与碳水化合物降解酶和底物的结合密切相关.虽然CBM不具有催化活性,但经证明CBM在纤维素[38]、木聚糖[39]、几丁质[40]和褐藻胶[41]等多糖降解酶结合底物的过程中起到关键作用.因此构建催化结构域+CBM的杂合酶,或在原有的基础上增加CBM个数都是提高酶结合转化底物的有效策略(表1).Duan等[42]将纤维素酶Um Ce19A与6种来自不同家族(CBM1、CBM2、CBM3、CBM4、CBM10和CBM72)的CBM融合构建杂合体蛋白,与野生型酶相比,融合了CBM的杂合酶对不溶性纤维素的催化活性和催化效率均有明显地提升,其中CBM4-Umcel9A较野生型对微晶纤维素的酶活提高6.6倍.将褐藻胶裂解酶Aly L2-FL通过与CBM13进行融合表达提高了其与可溶性褐藻酸盐的结合能力,从而提高了单催化模块(Aly L2-CM)的催化效率[41].来自嗜热纤维梭菌的内切纤维素酶Cc Cel9A由一个GH9催化结构域和C端5个CBM组成,通过对Cc Cel9A的CBM截短实验及结构分析,证明C端CBM3b和3个CBMx2使得该酶拥有了对纤维素的高效吸附能力,而GH9附近的CBM3c能与11个葡萄糖环紧密结合,从而将该酶的底物结合位点延伸到17个亚位点,大大增加了酶解效率[36].
| 图表编号 | XD00208342400 严禁用于非法目的 |
|---|---|
| 绘制时间 | 2021.03.20 |
| 作者 | 张舒、赵越、陈冠军、余俊红、吴秀芸、王禄山 |
| 绘制单位 | 山东大学微生物技术国家重点实验室、山东大学微生物技术国家重点实验室、山东大学微生物技术国家重点实验室、青岛啤酒股份有限公司啤酒生物发酵工程国家重点实验室、山东大学微生物技术国家重点实验室、山东大学微生物技术国家重点实验室 |
| 更多格式 | 高清、无水印(增值服务) |


{kind=link}