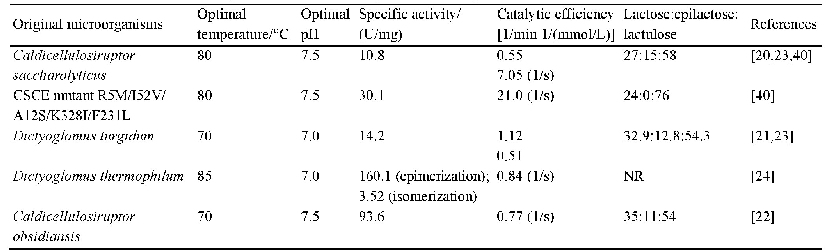
《表2 参与DON氧化和差向异构化的细菌及关键酶》
 提示:宽带有限、当前游客访问压缩模式
提示:宽带有限、当前游客访问压缩模式
本系列图表出处文件名:随高清版一同展现
《脱氧雪腐镰刀菌烯醇的生物转化及其隐蔽型毒素的形成研究进展》
注:+.表示具有该反应转化能力(数量与能力呈正相关);-.表示缺乏该反应转化能力;/.未知。
目前针对德沃斯式菌属的酶促差向异构化的研究更为深入,德沃斯式菌同属的不同菌株表现出来差向异构化的能力存在差异,这与菌株是否表达参与DON氧化及3-keto-DON还原的关键酶有关。Hassan等[54]将DON与7种Devosia属菌株一起孵育,发现只有D.mutans 17-2-E-8能将DON差向异构化为3-epi-DON,其他菌株没有显示出差向异构化DON的能力,但这几种Devosia菌株均能不同程度地将3-keto-DON转化为3-epi-DON。随后,同课题组的Carere等[55]从D.mutans 17-2-E-8中克隆了参与催化DON氧化形成3-keto-DON的基因,转化大肠杆菌进行表达,进一步测试了表达的降解酶Dep A活性及其转化DON的能力,发现该酶属于吡咯喹啉醌依赖的乙醇脱氢酶,接着用同样方法分离出参与异构化第二步反应的关键酶Dep B,该酶属于醛酮还原酶超家族[54]。最近,Wang Gang等[56]报道D.insulae A16对DON及其隐蔽型毒素3-Ac-DON和15-Ac-DON的最终产物降解为3-keto-DON,无法催化3-keto-DON转化为3-epi-DON,表明D.insulae A16缺乏特异性还原3-keto-DON的酶。这也进一步证实了DON差向异构化以3-keto-DON作为中间体,通过两步催化完成DON差向异构化(表2)。
| 图表编号 | XD00225605100 严禁用于非法目的 |
|---|---|
| 绘制时间 | 2020.10.15 |
| 作者 | 唐语谦、潘药银、刘晨迪、杨继国 |
| 绘制单位 | 华南理工大学食品科学与工程学院、华南协同创新研究院、华南协同创新研究院、华南理工大学食品科学与工程学院 |
| 更多格式 | 高清、无水印(增值服务) |


{kind=link}