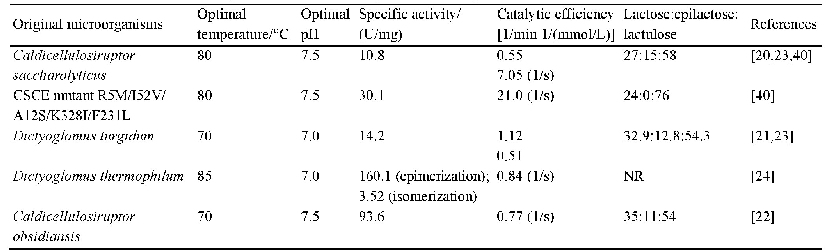
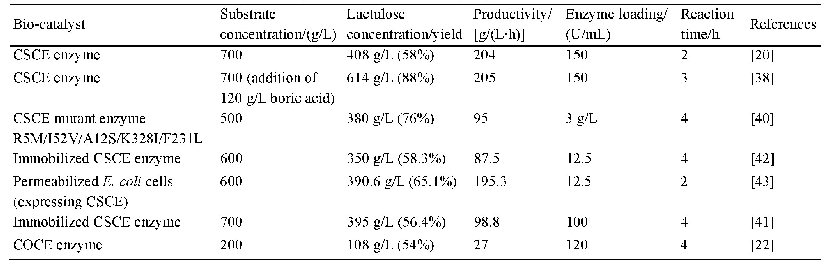
《表1.已报道催化乳糖能够产生乳果糖的纤维二糖差向异构酶》
 提示:宽带有限、当前游客访问压缩模式
提示:宽带有限、当前游客访问压缩模式
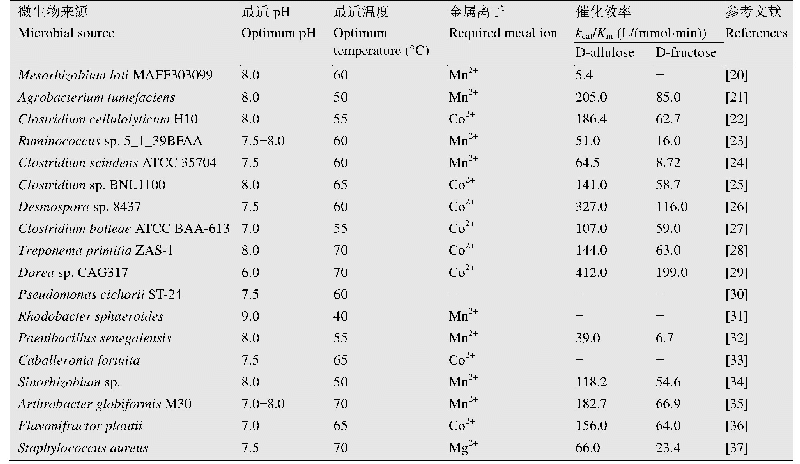
CE酶曾被认为是AGE酶(N-乙酰-D-氨基葡萄糖2-差向异构酶、EC 5.1.3.8)家族的成员,但由于AGE酶一般需要ATP参与反应,而CE酶并不需要任何辅酶,且两种酶的序列相似度很低,因此近年来将CE酶单独划为纤维二糖差向异构酶。早在1967年Tyler和Leatherwood就研究了革兰氏阳性菌Ruminococcus albus (白色瘤胃球菌)来源的CE酶,发现它可以催化纤维二糖产生葡萄糖基甘露糖[33],然而CE酶成为研究热点却是在40多年后。2011年韩国的Park等发现Caldicellulosiruptor saccharolyticus来源CE酶(CSCE)可以催化单糖D-葡萄糖产生D-甘露糖(同时产生副产物D-果糖)[34],由于D-甘露糖的应用价值,很快CE酶得到了重视。仅一年后通过继续研究,Kim和Oh又发现CSCE酶可以催化乳糖产生乳果糖[20],自此拉开了CE酶应用于乳果糖合成的序幕。乳糖的溶解度随温度变化很大,其中0°C下溶解度仅为11.9%(W/W),而74°C却达到86.2%(W/W)[35],这表明高的反应温度可以溶解更多的乳糖,从而提高产物浓度和生产强度。已报道可以产乳果糖的CE酶均具有较高的反应温度,在70–80°C;温度还可以影响CE酶催化醛酮糖异构(乳糖变为乳果糖)和差向异构(乳糖变为依匹乳糖)的产物比例,Park等发现CSCE酶在65°C下的醛酮糖异构活性是37°C下的12倍,而差向异构活性仅为37°C下的27%[36]。因此提高反应温度对高产乳果糖和降低依匹乳糖含量有很大帮助。由表1可知,目前已知的利用纤维二糖差向异构酶催化乳糖得到乳果糖的报道中,都会得到依匹乳糖这一反应副产物,包括Caldicellulosiruptor saccharolyticus、Dictyoglomus turgidum、Dictyoglomus thermophilum、Caldicellulosiruptor obsidiansis等来源(这里均指野生型酶),所产依匹乳糖的含量大约为11%–16%(W/W)[20–24]。依匹乳糖的结构与乳糖和乳果糖都非常相似,属于同分异构体,因此分离极为困难[37],目前还没有产业化规模下分离成功的报道。为了克服这一困难,Kim等提出在生物催化的过程中添加化学催化剂硼酸,可以使得依匹乳糖的含量降低到2%的水平[38]。其原理是硼酸可以和酮糖(乳果糖)形成络合物,使酮糖脱离反应体系,导致反应平衡向更多酮糖产物生成方向移动。由于依匹乳糖不含酮基,因此在硼酸体系中含量会大幅下降。硼酸的使用也见于D-塔格糖等稀少糖的催化研究中[39],然而硼酸的使用量较大,而硼是禁止出现在食品中的,因此必须完全去除才能符合法律标准。但作为一种弱酸,想要完全去除非常困难,即使价格昂贵的专用树脂也比较困难,因此这一方法的产业化价值有限。尽管许多糖基异构酶都被证明是金属酶,金属离子对CE酶的活性却没有促进作用,而且经EDTA处理活性也未发生变化,这表明CE酶并不是金属酶[34]。已报道的CE酶热稳定性大多较好,例如Dictyoglomus turgidum CE酶在70°C的活性半衰期达到了55 h[21]。
| 图表编号 | XD00212138400 严禁用于非法目的 |
|---|---|
| 绘制时间 | 2021.02.04 |
| 作者 | 徐铮、张倩、李克文、徐虹 |
| 绘制单位 | 南京工业大学食品与轻工学院材料化学工程国家重点实验室、保龄宝生物股份有限公司、保龄宝生物股份有限公司、南京工业大学食品与轻工学院材料化学工程国家重点实验室 |
| 更多格式 | 高清、无水印(增值服务) |

{kind=link}