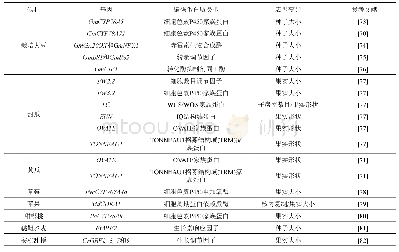
《表3 其他被子植物中参与果实/种子大小调控的基因》
 提示:宽带有限、当前游客访问压缩模式
提示:宽带有限、当前游客访问压缩模式
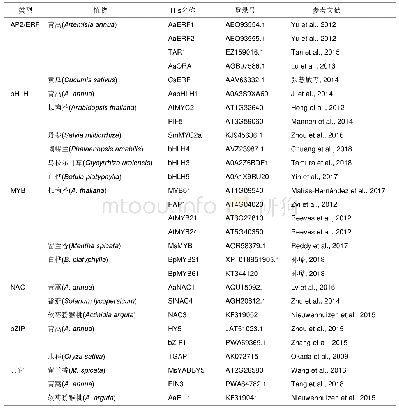
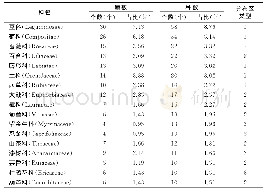
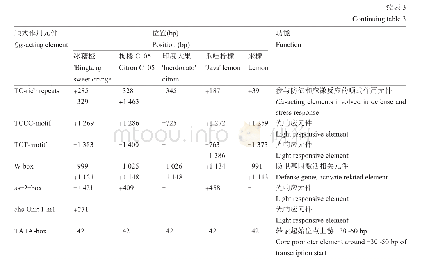
研究者也开始关注一些有经济价值的作物、蔬果植物,如大豆、葫芦科的黄瓜、甜瓜和罗汉果、蔷薇科的草莓、苹果、沙梨和樱桃、廖科的苦荞麦、胡颓子科的沙棘、葡萄科的葡萄和芸香科的柑橘等,对它们果实大小的遗传调控也有一些报道(表3).但是,在这些植物中,很难像模式植物一样进行大规模的正向遗传学研究,而主要还是通过反向遗传学,如同源克隆、组学分析等技术获得一些候选基因(表3).研究发现,在大豆中一些基因的大量表达,如细胞色素P450家族(cytochrome P450)基因GmCYP78A5和GmCY-P78A72,GIBBERELLIN 20 OXIDASE(GmGA20OX)和NUCLEAR TRANSCRIPTION FACTOR Y SUBUNIT A(GmNFYA)等,和一些基因表达的下调或沉默,如BIG SEEDS(GmBS1和GmBS2)和CELL WALL INVERTASE INHIBITORS(GmCIF1),均使种子增大[70,73~76].我们通过栽培大豆和野大豆果实发育转录组的比较也鉴定了一系列参与种子发育,进而可能调控种子大小的候选基因[83,84].对比番茄果实大小调控的QTL,在甜瓜的基因组中鉴定到FAS,OVATE,FW2.2,SUN,LC和FW3.2等相关基因家族成员[77].进一步研究发现,OVATE和TONNEAU1等QTL/基因控制甜瓜、黄瓜的果实形状和马铃薯块茎形态[71].基于组学(基因组、比较转录组和表达谱)技术的分析,发现黄瓜大小和形状的变化由受精前后纵向和横向的细胞分裂和细胞大小的差异引起[85];在罗汉果早期果实上调的基因中包括许多参与调控果实生长发育、果实大小和成熟代谢的基因[86].在草莓整个果实发育过程中,生长素、赤霉素(GA)和脱落酸(ABA)形成相互关联的反馈环,来控制果实发育过程中ABA的水平;草莓cytochrome P450 707A4a(FveCYP707A4a)基因作为中心环节,不仅调节ABA的积累,还通过整合生长素、GA和ABA信号来协调果实大小和成熟时间[78].嘎拉苹果自然突变体“Grand Gala”果实直径增加15%,重量增加38%,研究表明,其果实的增大是由果肉细胞变大引起的;进一步分析表明,MdCDKA1(cyclin dependent kinases 1)基因表达上调,细胞周期蛋白基因CYCA2;3表达下调,从而促进核内复制使细胞变大[79].在甜樱桃中发现,一个细胞色素P450基因PaCYP78A9(cytochrome P450 78A9)可以调控果实大小[80].比较转录组分析发现,沙梨果实发育早期与果实大小相关的差异表达基因中包含38个转录因子、23个细胞分裂素脱氢酶和赤霉素双加氧酶基因[87].在蓼科荞麦属苦荞麦的研究中发现,生长素和脱落酸的平衡可能是调控细胞分裂的关键因素,它们通过激素信号通路诱导生长素反应因子auxin response factor 2(FtARF2)的表达上调,对种子大小进行调节[81].对种子大小有明显差异的沙棘进行miRNA高通量测序分析,鉴定出137个已知miRNA和264个新miRNA;进一步预测发现22个种子大小相关基因[88].在葡萄中,利用图位克隆,发现6个与葡萄果重相关的QTL,平均贡献率在11.3%~33.0%[89].芸香科柑橘具有非呼吸跃变的果实成熟行为和独特的解剖学果实结构.通过对血橙“早红”和脐橙“二十一世纪”的果实进行比较转录组和蛋白质组学分析研究,发现在幼果、着色开始和果实延迟采收两个月这三个连续阶段,两个品种间具有相当多的差异表达的基因与蛋白,这些基因/蛋白分别参与果实的生长发育、花青素合成和可溶性固形物积累等方面[90].安柳甜橙(Citrus sinensis cv.Anliu)中Growth-regulating factor基因CsGRF2,3,7和9在果实细胞膨大中起重要作用,它们可能通过GA途径或GA非依赖性途径实现对细胞大小的调控[82].而且,茄科植物的OVATE,SUN,FAS等控制果实形态的调控因子在芸苔科植物中也是通过影响纵向或横向的细胞分裂,从而调控果实生长、大小和形状的,具有相当的保守性[91].
| 图表编号 | XD0034730900 严禁用于非法目的 |
|---|---|
| 绘制时间 | 2019.04.20 |
| 作者 | 贺超英、王丽、严立新、李巧茹、雍斌、朱韦韦 |
| 绘制单位 | 中国科学院植物研究所系统与进化植物学国家重点实验室、中国科学院大学、中国科学院种子创新研究院、中国科学院植物研究所系统与进化植物学国家重点实验室、中国科学院植物研究所系统与进化植物学国家重点实验室、中国科学院大学、中国科学院植物研究所系统与进化植物学国家重点实验室、中国科学院大学、中国科学院植物研究所系统与进化植物学国家重点实验室、中国科学院大学、中国科学院植物研究所系统与进化植物学国家重点实验室、中国科学院大学 |
| 更多格式 | 高清、无水印(增值服务) |



{kind=link}