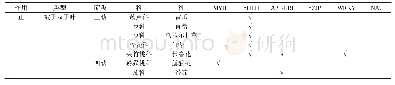
《表1 植物中调控落花落果的部分基因》
 提示:宽带有限、当前游客访问压缩模式
提示:宽带有限、当前游客访问压缩模式
注:影响落花落果的基因大致有SHATTERING类基因和MADS-box基因。SHATTERING类基因普遍存在于粮食作物(水稻、高粱、玉米等)和模式植物(拟南芥等)中,这类基因主要以调控离层细胞的形成和发育为主。MADS-box基因在大豆、拟南芥等荚果类植物中广泛存在,这些基因主要被划分为
大豆裂荚与器官脱落相似,其QTL也不容忽视,大豆裂荚的QTL已取得巨大进步,直至2014年,Suzuki et al(2010)、Dong et al(2014)和Funatsuki(2014)已报道了果荚开裂基因Pdh1和5个离区形成基因SHAT1-5,如图1所示。最早被定位于16染色体上的果荚开裂基因为Pdh1,来源于抗裂荚品种和易裂荚品种PI 416937的重组自交系群体,基因Phd1(Gm16 g25580)位于47 kb基因组区域(29,621-29,668 kb)的上游20 kb处,是造成大豆裂荚的基因,主要由于单核苷酸突变(A/T)导致提前终止密码子的出现,进而调节果荚开裂的大小。Pdh1基因编码同源蛋白,并在富含木质素的果荚壁内的厚壁组织中高度表达,所以说基因Phd1能调节木质素合成和果荚的开裂。此外,Yamada et al(2009)以种群抗裂荚品系Harosoy×易裂荚品系Toyomu-sume F2和Kariyutaka×Wasekogane F2两个群体为材料,把裂荚相关QTL定位于qPDH1附近并解释了抗裂荚基因尽管遗传背景有所不同,但QTL也定位于qPDH1附近,产生这种结果,最有可能是亲本为杂合基因,子代表现出高裂荚性状所致,裂荚抗性性状表现为近隐性性状,主效QTL在q PDH1处。Dong et al(2014)通过比较野生型和栽培型大豆之间的核苷酸遗传多样性,其中两个基因Gm04g39210和Gm16 g02200在驯化过程中核苷酸多样性降低,而且大豆种质在两个候选基因中没有遗传变异,Gm16g02200与大豆荚果开裂相关的QTL重叠。遗传和功能分析表明,被名为SHATTERING1-5(SHAT1-5)的Gm16g02200是拟南芥At NST1/2的直系同源基因,编码NAC结构域转录因子(表1)。基因表达和转基因互补分析的实验证实:SHAT1-5通过增加其在纤维帽细胞(Fiber Cap Cells,FCC)中的自身表达来控制栽培大豆中的抗裂荚表型。SHAT1-5和Pdh1共同调节作物品种中的次生细胞壁增厚和抗落粒性(Dong et al,2014)。4.3.4荞麦在蓼科荞麦属植物中,岳鹏等(2012)将SSR引物成功用于甜荞遗传图谱的构建和种质资源和农艺性状(如落粒性)研究,通过遗传作图表明:落粒基因Sht1和Sht2分别与标记S1182-1160和S1182-1048紧密连锁,Sht1和Sht2分子调控功能尚不清楚,其他落粒基因也未被发现,可以说荞麦属植物落粒性的研究相对匮乏,需要进一步努力。
| 图表编号 | XD0022855900 严禁用于非法目的 |
|---|---|
| 绘制时间 | 2018.09.01 |
| 作者 | 崔娅松、陈庆富、霍冬敖、李洪有 |
| 绘制单位 | 贵州师范大学荞麦产业技术研究中心、贵州师范大学荞麦产业技术研究中心、贵州师范大学荞麦产业技术研究中心、贵州师范大学荞麦产业技术研究中心 |
| 更多格式 | 高清、无水印(增值服务) |





{kind=link}