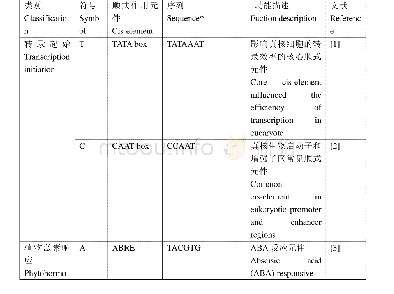
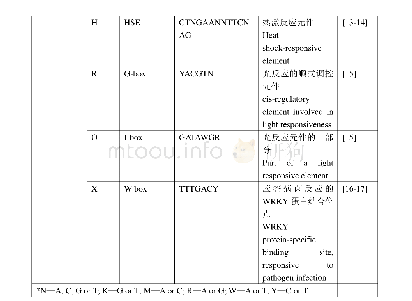
《附表CLF参与调控的基因》
 提示:宽带有限、当前游客访问压缩模式
提示:宽带有限、当前游客访问压缩模式
本系列图表出处文件名:随高清版一同展现
《表观遗传调控基因CLF在植物器官形态建成的研究进展》
除了本文上述所讨论的基因外,还有许多其他基因也受到CLF的调控。在野生型拟南芥中,至少有6 854个以上的基因位点是H3K27三甲基化的富集区域,这些靶基因受到PRC2蛋白调控,往往具有时空表达特异性[35]。如I类KNOX(KNOTTED1-LIKE HOMEOBOX)基因,它能够在胚胎发生和整个成熟发育过程中在茎顶端分生组织中起作用,维持细胞分裂不分化的状态[36,37]。DOG1(DELAYOF GERMINATION1)据报道是种子休眠的主要调节基因,它的缺失会形成不休眠种子[38-40]。DOG1在种子休眠前成熟过程中表达,其积累与休眠水平相关。低温也会上调DOG1的表达从而抑制休眠被打破[39]。而CLF表达会增加DOG1位点上H3K27三甲基化沉积,负调控DOG1,打破休眠并诱导萌发[41]。由CLF和SWN控制的H3K27三甲基化在植物脱落酸诱导的响应元件上富集,有助于长期抑制ABA诱导的衰老,并潜在地加强了植物的胁迫耐受性[42]。FIT(FER-LIKE IRON DEFICIENCY IN-DUCED TRANSCRIPTION FACTOR)推测是铁元素摄取反应的转录因子,CLF突变后,FIT表达量升高,研究发现clf突变体比野生型更耐铁缺乏[43]。AGO1(ARGONAUTE1)可以选择性招募micro RNA和siRNA并切碎他们。CLF突变后,由于AGO1泛素化增加导致特定组织的miRNA活性因此受损[44]。由此可知,CLF在植物整个生长发育和形态建成中起到了十分重要的作用。附表为CLF参与调控的基因。
| 图表编号 | XD00148866000 严禁用于非法目的 |
|---|---|
| 绘制时间 | 2020.06.25 |
| 作者 | 姜晓旭、李晓屿、马瑶、Ralf Müller-Xing、邢倩 |
| 绘制单位 | 东北林业大学、东北林业大学盐碱地生物资源环境研究中心、东北林业大学生命科学学院、辽宁石油化工大学化学化工与环境学部、东北林业大学、东北林业大学盐碱地生物资源环境研究中心、东北林业大学生命科学学院、东北林业大学、东北林业大学盐碱地生物资源环境研究中心、东北林业大学生命科学学院、东北林业大学、东北林业大学盐碱地生物资源环境研究中心、东北林业大学生命科学学院 |
| 更多格式 | 高清、无水印(增值服务) |




{kind=link}