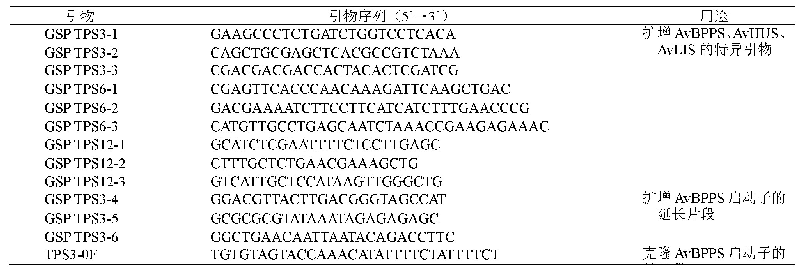
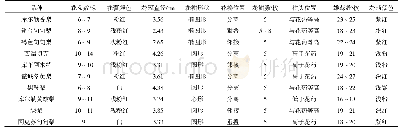
《表2 花器官特异性启动子》
 提示:宽带有限、当前游客访问压缩模式
提示:宽带有限、当前游客访问压缩模式
在花粉组织特异性启动子的发掘上,Peng等[18]研究证实Zm908启动子是花粉特异性启动子,-126~+68 bp区域对于花粉表达至关重要,5'端缺失分析显示-126~102 bp区域为转录抑制元件。Zm Dof30转录因子在体内和体外均能与Zm908启动子中AAAG元件(-126~+58 bp区)结合,并负向调控转基因烟草叶片中的p184区域活性。此外,玉米丝氨酸/苏氨酸激酶基因ZmSTK2 USP启动子序列包含花药/花粉特异性元件(GTGA和AGAAA),组织染色表明该启动子驱动GUS在花粉发育中期至成熟期特异性表达[19]。水稻转基因研究表明,启动子PLP1、PLP2(pollen late stage promoter)仅在水稻晚期花粉粒中驱动GUS表达。突变分析显示,PLP2中-740~-557 bp区域AGAAA和CAAT元件起增强子的作用。功能验证结果表明,两个启动子核心区域的TA富集区(TACATAA和TATTCAT)对于水稻花粉特异性表达是必要的[20]。POLLEN1LELAT52和GTGA元件为pZm908、ZmSTK2_USP和PLP2三个启动子共有元件(表2),决定着启动子花粉组织特异性表达,为发掘或人工构建新型花粉特异启动子提供理论支撑。
| 图表编号 | XD0038501700 严禁用于非法目的 |
|---|---|
| 绘制时间 | 2019.01.01 |
| 作者 | 焦勇、柳小庆、江海洋、陈茹梅 |
| 绘制单位 | 安徽农业大学生命科学学院、中国农业科学院生物技术研究所、中国农业科学院生物技术研究所、安徽农业大学生命科学学院、中国农业科学院生物技术研究所 |
| 更多格式 | 高清、无水印(增值服务) |




{kind=link}