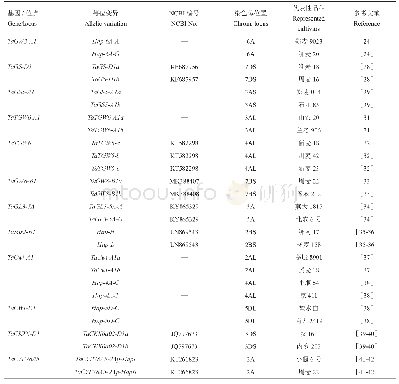
《表1 小麦中已发现的粒重形成相关基因及其变异类型》
 提示:宽带有限、当前游客访问压缩模式
提示:宽带有限、当前游客访问压缩模式
本系列图表出处文件名:随高清版一同展现
《小麦粒重相关基因的遗传定位和分子标记辅助育种进展》
小麦属于异源六倍体、基因组庞大、重复序列较多,给克隆和分离小麦基因带来很大困难,但是随着小麦全基因组序列信息的公布及现代分子克隆技术的快速发展,小麦粒重形成相关基因的克隆也取得较快发展。Su等[24]利用控制水稻粒宽和粒重的OsGW2基因序列,同源克隆得到小麦Ta GW2基因,该基因编码区全长1275 bp,编码424个氨基酸。同时,利用中国春的缺体-四体系将其定位于第6同源群上,并在TaGW2-6A基因的启动子区域发现了2个SNP位点,分别命名为Hap-6A-A和Hap-6A-G,同时发现Hap-6A-A类型在小麦籽粒发育过程中发挥了更为积极的作用。但之后,Zhang等[25]研究发现TaGW2-6A基因在粒宽、粒长和千粒重方面呈下调表达,认为Hap-6A-G单体型是控制小麦粒重的优异等位基因。Yang等[26]对TaGW2基因进一步研究发现该基因含有8个外显子和7个内含子以及1个保守的功能结构域,并在兰考大粒中发现在6A染色体上TaGW2基因的1个等位变异,即在第8个外显子上977 bp处有1个单碱基T的插入,从而使该基因翻译提前终止。同时发现该突变导致小麦粒宽和千粒重显著增加,推测认为TaGW2基因对小麦粒重的发育起负向调控作用[26]。Simmonds等[27]利用TILLING技术在普通小麦中发现TaGW2-A1基因的1个新突变类型,即在第5外显子处由G变A导致发生错误剪接,且发现GW2-A1突变体与野生型相比能显著增加千粒重(6.6%)、粒宽(2.8%)、粒长(2.1%)。Zhang等[28]利用同源克隆的方法从普通小麦7DS染色体上克隆得到TaGS-D1基因,其包含4个外显子和3个内含子,cDNA全长255 bp,编码85个氨基酸。在不同小麦品种中扩增TaGS-D1基因发现其含有2种等位变异,分别命名为TaGS-D1a和TaGS-D1b,且在拥有不同等位变异类型品种的千粒重间存在显著性差异。Wang等[29]利用同源克隆技术从普通小麦中克隆了一个控制籽粒大小的TaGS5-A1基因,该基因由10个外显子和9个内含子组成,被定位在3AS染色体上,并发现了其至少存在2种等位变异,分别命名为TaGS5-A1a和TaGS5-A1b。之后,在TaGS5-A1基因启动子上游顺式作用元件Sp1内发现单碱基G的插入,并将先前在TaGS5-A1基因第6个外显子内发现的SNP(G/T)导致的等位变异类型和其启动子区域发现的G碱基插入与否而造成的等位变异类型相结合,把普通小麦又分为TaGS5-A1a-a、TaGS5-A1a-b、TaGS5-A1b-a和TaGS5-A1b-b 4种基因型,进一步研究发现,拥有TaGS5-A1a-b和TaGS5-A1b-b类型的小麦品种具有较宽的粒宽和更高的千粒重[30]。Hanif等[31]和Hu等[32]利用水稻中的TGW6基因序列进行同源克隆,其中Hanif等[31]克隆得到位于普通小麦3AL染色体上的TaTGW6-A1基因,含有987 bp的开放阅读框,并发现该基因在不同品种中存在着单核苷酸多态性,形成2个单倍型,分别命名为TaTGW6-A1a和TaTGW6-A1b。而Hu等[32]则克隆得到TaTGW6基因,该基因全长1656 bp,仅包含1个外显子和1个含有1035 bp的开放阅读框,并将其定位在普通小麦4AL染色体上,且在TaTGW6位点发现3种等位变异类型,分别命名为TaTGW6-a、TaTGW6-b和TaTGW6-c,进一步研究发现TaTGW6-b和TaTGW6-c变异类型的品种在粒径和粒重方面显著高于TaTGW6-a类型,但TaTGW6-b和TaTGW6-c变异类型在当前品种中分布概率较低。最近,Yan等[33]在矮抗58中克隆了控制粒重的基因,命名为TaGW8,被定位于7BS染色体上,而Yang等[34]利用水稻GL3基因序列同源克隆出控制小麦粒长和粒重的TaGL3-5A基因。通过对不同籽粒大小的小麦品种基因多态性分析,发现TaGW8-B1基因存在2种变异类型,即第1个内含子有一个276 bp的InDel,有276 bp InDel命名为TaGW8-B1b,没有的则为TaGW8-B1a[33]。TaGL3-5A基因全长10227bp,由21个外显子和20个内含子组成,被定位在5A染色体上,根据其单核苷酸多态性发现其有2种等位变异,分别命名为Ta GL3-5A-A和Ta GL3-5A-G[34]。与此同时,小麦体内一些对粒重形成和发育至关重要酶类物质及其相应的基因也相继被定位和克隆。Jiang等[35]在普通小麦的2A、2B和2D染色体上发现了蔗糖合成酶基因,分别为TaSus2-2A、TaSus2-2B和TaSus2-2D,且在TaSus2-2B基因上发现3个单核苷酸多态性,形成2个单倍型,分别命名为Hap-H和Hap-L。之后,Volpicella等[36]在硬粒小麦中也克隆得到Sus2-2B基因,并对其功能进行研究。小麦体内的细胞壁结合转化酶在碳水化合物的转运中发挥着重要作用。Ma等[37]在普通小麦的2AL染色体上克隆得到细胞壁转化酶TaCwi-A1基因,该基因全长3676 bp,含有7个外显子和6个内含子以及1个含有1767 bp的开放阅读框,发现该基因的2种等位变异,分别命名为TaCwi-A1a和TaCwi-A1b,进一步研究发现TaCwiA1a变异类型与高千粒重的品种相关,而TaCwiA1b类型则与低千粒重的品种相关。之后,Jiang等[38]在普通小麦4A、5B和5D染色体上克隆得到TaCWI基因,其中TaCWI-4A和TaCWI-5D基因也都发现了相应的单体型,分别为Hap-4A-C、Hap-4A-T和Hap-5D-G、Hap-5D-C,进一步研究发现拥有Hap-4A-T和Hap-5D-C类型的品种千粒重显著高于Hap-4A-C和Hap-5D-G类型的品种。细胞分裂素氧化酶在植物生长和发育过程中扮演极其重要的角色。在普通小麦中克隆得到细胞分裂素氧化酶TaCKX6-D1基因,该基因隶属于TaCKX基因家族,被定位在3DS染色体上[39],随后,Lu等[40]发现在TaCKX6a02位点发现2种等位变异类型,分别命名为TaCKX6a02-D1a和TaCKX6a02-D1b。细胞色素P450(CYP,Cytochromes P450,)家族是最大的植物蛋白家族之一,其中CYP78A(cytochromes P450 78A)家族的基因可应用于作物改良。Ma等[41]利用同源克隆的方法获得小麦TaCYP78A5基因,并利用瞬时沉默和过表达技术揭示TaCYP78A5表达水平与小麦粒重呈正相关。之后,司文洁等[42]又根据不同品种小麦TaCYP78A5启动子区序列内SNP位点差异开发TaCYP78A5-2A启动子区功能标记CAPS-5Ap,并对其进行验证。此外,小麦体内的果聚糖合成酶(6-SFT-A2)、可溶性淀粉合成酶(SSS)、腺苷二磷酸葡萄糖焦磷酸化酶(AGPP)等是小麦粒重形成过程中的关键酶,对小麦粒重的提高具有重要的调节作用[43-44]。吕广德等[45]对普通小麦粒重相关基因克隆的研究进展进行了详细的综述。截止目前,已有多个小麦粒重形成相关基因及其变异类型在普通小麦中被发现克隆,详细信息见表1。
| 图表编号 | XD00181531400 严禁用于非法目的 |
|---|---|
| 绘制时间 | 2020.05.16 |
| 作者 | 张福彦、范家霖、陈晓杰、陈锋、齐红志、王嘉欢、程仲杰、杨保安、张建伟 |
| 绘制单位 | 河南省科学院同位素研究所有限责任公司、河南省核农学重点实验室、河南省科学院同位素研究所有限责任公司、河南省核农学重点实验室、河南省科学院同位素研究所有限责任公司、河南省核农学重点实验室、河南农业大学农学院、河南省粮食作物协同创新中心、河南农业科学院农业经济与信息研究所、河南省科学院同位素研究所有限责任公司、河南省核农学重点实验室、河南省科学院同位素研究所有限责任公司、河南省核农学重点实验室、河南省科学院同位素研究所有限责任公司、河南省核农学重点实验室、河南省科学院同位素研究所有限责任公司、河南省核农学重点 |
| 更多格式 | 高清、无水印(增值服务) |




{kind=link}