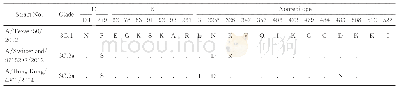
《表3 2014~2018年临沧市H3N2亚型流感病毒HA蛋白氨基酸突变位点分析》
 提示:宽带有限、当前游客访问压缩模式
提示:宽带有限、当前游客访问压缩模式
本系列图表出处文件名:随高清版一同展现
《新一代全基因组测序揭示2014~2018年云南省临沧市H3N2亚型流感病毒的遗传突变机制》
注:氨基酸比对参照A/Texas/50/2012,“a”表示受体结合位点,“.”为氨基酸位点与A/Texas/50/2012对应位点相同。
29株H3N2亚型流感病毒HA蛋白的裂解位点序列为PEKQTR↓G或PERQTR↓G,均含有碱性氨基酸R,属于低致病性流感病毒。与WHO推荐的北半球2014~2015年度疫苗株A/Texas/50/2012的HA氨基酸序列相比,29株H3N2流感病毒HA蛋白共发生42处位点变异(不包含信号肽),32处位于HA1,10处位于HA2区,分别占76.2%与23.8%(表2和表3)。HA蛋白共有5个抗原决定簇(A-E),全部位于HA1亚基[11],29株病毒株在5个抗原决定簇上共有29处位点发生突变,位于抗原表位A区的N145S、B区的V186G和D区的F219S已形成适应性变异。突变位点统计分析发现归属3C.2a1进化分支的11株毒株几乎都发生了L3I(10/11株)、N128T(10/11株)、F159Y、K160T和D489N变异,其中部分毒株还发生了K92R(7/11株)、N121K(8/11株)、T135K(7/11株)、N144S(8/11株)、N171K(7/11株)、Q311H(4/11株)、I406V(7/11株)、G484E(7/11株)变异,已有文献报道N121K、N171K、I406V和G484E这四个位点变异具有3C.2a1新进化分支的专属性[12],所以这很好地解释了为什么这11株毒株并入3C.2a1进化簇。A/Yunnan-Linxiang/114/2018毒株独具K83R、N122D、L157S、Q197H和D225N变异,并入3C.3b进化分支[10]。从表2和表3还可以看出HA蛋白N128A、A138S、F159S、K326R突变是3C.3a进化分支的代表性突变位点,这与相关文献报道一致[13]。隶属3C.2a1进化分支的2016~2018年10株毒株还在受体结合位点134~138残基处[14]发生了T131K或T135K/N(A区)变异,这与归属3C.3a进化分支的17株毒株发生在抗原决定簇A138S(A区)的突变同属RBS突变。有研究报道流感病毒HA突变位点如同时属于抗原决定簇和RBS及其附近位点,则被认为是抗原漂变的决定位点[11]。
| 图表编号 | XD00144512800 严禁用于非法目的 |
|---|---|
| 绘制时间 | 2020.09.01 |
| 作者 | 张汉菊、仇娜、周晓荣、赵晓南、徐闻、伏晓庆、张美玲 |
| 绘制单位 | 临沧市疾病预防控制中心、临沧市疾病预防控制中心、临沧市疾病预防控制中心、云南省疾病预防控制中心、云南省疾病预防控制中心、云南省疾病预防控制中心、云南省疾病预防控制中心 |
| 更多格式 | 高清、无水印(增值服务) |




{kind=link}