《表2 出发菌株M-Z18与SS-19菌株的关键酶酶活性比较(U/mg prot)》
 提示:宽带有限、当前游客访问压缩模式
提示:宽带有限、当前游客访问压缩模式
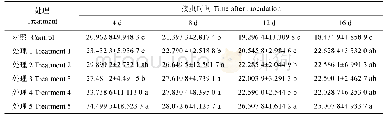
为了从生理代谢水平进一步了解SS-19的高产原因,对ε-PL合成途径的关键酶活性进行了考察,结果如表2所示。与M-Z18相比,SS-19的HK、G6PDH、PK、CS、PEPC、ASK、Pls的酶活性分别增加了40.23%、45.45%、28.13%、142.11%、119.27%、136.49%、34.79%。HK酶活升高会增加EMP途径通量,有助于SS-19对葡萄糖的摄取和消耗。NADPH是ε-PL前体L-赖氨酸合成中最重要的辅因子,其主要来源于PPP途径。G6PDH酶活性提高,PPP途径会被强化,促使细胞合成更多的NADPH,从而促进了L-赖氨酸合成。EMP途径代谢末端产物丙酮酸是TCA循环所需的碳骨架,而关键酶PK的活力增强直接增加了碳骨架进入TCA循环的通量。同时,CS酶活力的增强也促使TCA循环的通量增加。TCA循环能为细胞提供能量因子(NADH和FADH2)用于呼吸产能,并生成大量ATP,直接参与ε-PL的合成代谢。在ε-PL合成中大量的L-赖氨酸会被消耗,细胞胞内草酰乙酸是相对稀缺的,PEPC酶活性增加提高了回补途径的通量,为前体L-赖氨酸合成提供大量的碳骨架前体(草酰乙酸)并流入DAP途径。ASK酶活性的提高会强化DAP途径,促进L-赖氨酸的合成。此外,Pls的活力的增加直接提高了细胞的ε-PL合成能力。以上结果表明,SS-19是通过强化初级与次级代谢通量,合成更多前体物质L-赖氨酸和辅因子ATP,最终大量合成ε-PL的。这与许召贤等将VHb基因整合到S.albulus PD-1的染色体中以减少氧限制并增强发酵中ε-PL的生物合成和Xu等通过在S.albulus PD-1中过表达基因amt B来改善ε-PL的产量的结果是一致的[8,9]。
| 图表编号 | XD00157510600 严禁用于非法目的 |
|---|---|
| 绘制时间 | 2020.03.20 |
| 作者 | 李芹、赵俊杰、刘永娟、王靓、毛忠贵、陈旭升 |
| 绘制单位 | 工业生物技术教育部重点实验室江南大学生物工程学院、工业生物技术教育部重点实验室江南大学生物工程学院、工业生物技术教育部重点实验室江南大学生物工程学院、工业生物技术教育部重点实验室江南大学生物工程学院、工业生物技术教育部重点实验室江南大学生物工程学院、工业生物技术教育部重点实验室江南大学生物工程学院 |
| 更多格式 | 高清、无水印(增值服务) |





{kind=link}