《表2 Rieske型加氧酶催化多样性》
 提示:宽带有限、当前游客访问压缩模式
提示:宽带有限、当前游客访问压缩模式
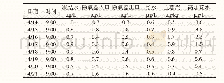
Rieske型芳香环双加氧酶能够催化多种反应,包括羟基化、环氧化、去饱和、氯化、脱烷基、去甲基化等(图1)[14]。例如,萘双加氧酶不仅能够催化多种双环和三环的芳香族化合物双羟化反应生成对应的二氢二醇,而且能催化其他氧化反应,包括单羟化、去饱和、脱烷基化和亚砜化等[20]。同属非血红素铁依赖型的Fe2+/α酮酸双加氧酶也能够以加氧的方式催化类黄酮进行环羟化、氧化、去饱和等反应[21](表2)。Rieske型芳香环双加氧酶具有宽泛的底物特异性,Baboshin等[22]研究认为芳香环羟化双加氧酶催化芳香烃化合物的特异性存在两个特征:1)同一的双加氧酶可催化不同的底物羟基化;2)对于不同的底物,同一双加氧酶表现的活性不同。计算分析结果显示,来自Sphingobium sp.PNB的Rieske型芳香环双加氧酶具有降解多种芳香族化合物的能力,证明该加氧酶具有宽泛的底物特异性[10]。不同来源的联苯双加氧酶(BPDO)对同类底物的偏好和区域特异性存在显著差异,可以催化甲苯、多氯联苯、羟基联苯等底物[17,23]。来自Pseudomonas KF707的联苯双加氧酶BPDOKF707不能催化联苯的3,4-位双羟基化,但对二氯联苯同系物却表现出不同的活性,较2,2′-或3,3′-二氯联苯而言,优先转化4,4′-二氯联苯[24]。另外,虽然Burkholderia xenovorans LB400的联苯双加氧酶BPDOLB400与BPDOKF707的序列同源性超过95%,但BPDOLB400的转化范围更广。BPDOLB400能够优先转化含有多达6个氯的邻位取代同系物,也能催化2,2′,5,5′-四氯联苯3,4双羟基化,还能催化某些二氯同系物的脱卤作用,产生2,3-二羟基联苯。BP-DOLB400与Pandoraea pnomenusa B-356的联苯双加氧酶BPDOB-356均能催化联苯,但BPDOB-356具有更高的转化效率。联苯双加氧酶的α亚基(BphA1)直接影响酶的催化活性和底物特异性[25],该差异性是由α亚基上少量的氨基酸差异造成的[26]。BphA1活性位点中的氨基酸残基具有不同的作用,协同决定了酶的底物特异性[27]。β亚基对底物特异性的影响存在着两种不同的看法。Hurtubise等[28]通过对BPDOB-356和BPDOLB400的αβ亚基进行交叉重组实验,证明α和β亚基均影响酶与底物的相互作用,其中β亚基的结构不仅能影响酶对催化底物的氧结合能力,还能影响底物的反应模式。α和β亚基的结构特征决定了催化口袋的大小和形状,进而影响氧化底物的范围,如Sphingobium sp.PNB的α亚基结构包含一个非常大的底物结合口袋,可以满足容纳高分子量底物的空间要求,因此能够催化多种芳香烃化合物[10]。而Kauppi等[29]研究工作表明α亚基是控制底物特异性的主要成分,β亚基似乎不参与底物的催化,可能仅作用于酶的结构稳定性。
| 图表编号 | XD00186069800 严禁用于非法目的 |
|---|---|
| 绘制时间 | 2020.08.01 |
| 作者 | 刘玉、杨键、李茹、龙丽娟 |
| 绘制单位 | 中国科学院南海海洋研究所中国科学院热带海洋生物资源与生态重点实验室、中国科学院大学、中国科学院南海海洋研究所中国科学院热带海洋生物资源与生态重点实验室、南方海洋科学与工程广东省实验室(广州)、中国科学院南海海洋研究所中国科学院热带海洋生物资源与生态重点实验室、中国科学院大学、中国科学院南海海洋研究所中国科学院热带海洋生物资源与生态重点实验室、中国科学院大学、南方海洋科学与工程广东省实验室(广州) |
| 更多格式 | 高清、无水印(增值服务) |




![表1 Rieske型芳香环双加氧酶分类[16,19]](http://bookimg.mtoou.info/tubiao/gif/GXKX202003009_00600.gif)
{kind=link}