《表1 16S rRNA甲基化酶相关基因》
 提示:宽带有限、当前游客访问压缩模式
提示:宽带有限、当前游客访问压缩模式
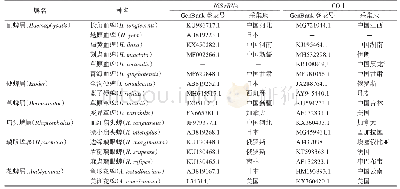
现已发现10组甲基化酶:Arm A、Rmt A-RmtH、NpmA[8],见图1。2002年首次检测出16S rRNA甲基化酶[9],2003年,Galimand等[5]发现其为ArmA。armA基因多位于可转移质粒的复合转座子上,后发现也可存在IS26或ISAba125复合转座子相关的独立环状结构,与宿主质粒共存[10],仍不明确其确切起源。同年发现的RmtA,其基因位于接合型质粒中的耐汞转座子Tn5041上,在移动元件κ-γ的两个拷贝之间,该基因可能源于产氨基糖苷放线菌[11]。2004年,Doi等[12]发现RmtB,其基因与rmtA核酸序列相似性高。ArmA和RmtB是世界范围报道最多的16S rRNA甲基化酶,虽然两者都易携带blaCTX-M,但rmtB与blaCTX-M位于不同的质粒[13]。2006年在奇异变形杆菌中发现RmtC[14],与之前发现的16S rRNA甲基化酶相似性不高。RmtC的检出率在一些地区仅次于ArmA和RmtB[15]。2007年,在巴西分离出同时产SPM-1金属酶和RmtD的细菌[16],在几年后又发现了rmtD2[17]与rmtD3[18]基因型。现RmtD相关报道主要集中于巴西,且有一定概率同时产SPM-1[19]。2010年Rmt E首次分离自牛源性大肠埃希菌[20],后在2013年于人体分离出[21],5年后发现rmt E2基因型[22],至今相关报道依旧十分罕见。2011年,在法国发现了RmtF,与RmtD编码蛋白一致性较高[23-24],其可能与放线菌的16S rRNA甲基化酶存在亲缘关系[25]。2011年在巴西筛选出5株产RmtG细菌,这5株均同时产CTX-M,有4株产KPC-2[26]。早先Rmt G的报道主要在美洲[27],2018年首次在欧洲出现[28]。2年后检测出RmtH[29],该菌株同时携带blaCTX-M-15、blaSHV-1、blaOXA-1[30]。2007年发现了卡那霉素-阿泊拉霉素甲基转移酶(Kam)家族的NpmA[2]。NpmA可能拥有m(1)A1408和m(1)G1408的双重活性[31]。现在产NpmA的菌株依旧较为罕见。现已发现16S rRNA甲基化酶基因的相关研究见表1[32-45]。
| 图表编号 | XD004577800 严禁用于非法目的 |
|---|---|
| 绘制时间 | 2019.11.20 |
| 作者 | 施清喻、胡付品 |
| 绘制单位 | 复旦大学附属华山医院抗生素研究所国家卫健委临床药理重点实验室、复旦大学附属华山医院抗生素研究所国家卫健委临床药理重点实验室 |
| 更多格式 | 高清、无水印(增值服务) |


{kind=link}