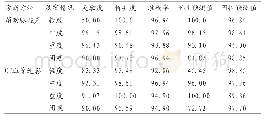
《表2 寒温带和热带地区鸟类基础代谢率(BMR)实测值与预测值的比较》
 提示:宽带有限、当前游客访问压缩模式
提示:宽带有限、当前游客访问压缩模式
BMR是TNZ内动物维持正常生理功能所需的最小产热率,是反映不同物种和个体间能量消耗水平的重要生理指标(Mc Kechnie,2008;Swanson et al.,2017)。北红尾鸲的BMR是体重预期值的151%,明显高于热带地区鸟类的体重预期值(平均为83%)(Weathers,1979),表现出寒温带鸟类的代谢特点。Weathers(1979)和Mc Nab(2009)认为,鸟类BMR与气候特点密切相关,生活在低纬度地区小型鸟类的BMR一般较预期值要低,而生活在高纬度地区小型鸟类的BMR一般要高于预期值。根据Londo1o等(2015)的异速公式,部分发表的热带鸟类和寒温带BMR数据及体重预期值见表2。如生活在温暖的环境,鸟类较低的BMR反映出较低的能量消耗,与较慢的生活节律(slow pace of life)和较低的繁殖投入有关(Wiersma et al.,2007)。如斑胸草雀(Poephila guttata)主要分布于澳大利亚和印度尼西亚的干旱环境,降低代谢产热是其为减少水分蒸发,保持水分的一种适应方式(Marschall et al.,1 991)。橙颊梅花雀(Estrilda melpoda)分布于东非的塞内加尔、甘比亚和刚果等地;辉胸闪羽蜂鸟(Aglaeactis castelnaudii)、蓝翅大蜂鸟(Pterophanes cyanopterus)和白腹林蜂鸟(Chaetocercus mulsant)主要分布在中南美及加勒比海地区。这些地区全年气温较高,湿度大,四季界限不明显,降低代谢产热及减少繁殖投入是有效的适应特征(Londo1o et al.,2015)。相反,生活在寒冷地区鸟类由于自身维持的能量代价相对较高,繁殖季节短暂,高的BMR是对低温和高繁殖投入的适应(Liu et al.,2004)。如鸲岩鹨(Prunella rubeculoides)、棕颈雪雀(Montifringilla ruficollis)和黄嘴朱顶雀(Acanthis flavirostris)BMR的体重预期值相应为237%、211%和227%(邓合黎等,1990);北朱雀(Carpodacus roseus)、燕雀(Fringilla montifringilla)和白腰朱顶雀(Acanthis flammea) BMR的体重预期值分别为242%、222%和210%(Liu et al.,2004);太平鸟(Bombycilla garrulous)和灰头鹀(Emberiza spodocephala) BMR的预期值相应为175%和239%(李铭等,2005)(表2)。如鸲岩鹨和黄嘴朱顶雀分布于我国青藏高原,其热调节有明显的高寒草甸动物代谢的特征(邓合黎等,1990);北朱雀、燕雀、白腰朱顶雀、太平鸟和灰头鹀繁殖于亚洲东部及西伯利亚等地,具有北方动物代谢的特点(Liu et al.,2004;李铭等,2005);而小鹀(E.pusilla)表现迁徙性鸟类的特点,繁殖地高于越冬地及中途停歇地,在我国南方(温州)和北方(齐齐哈尔) BMR的体重预期值相应为177%和219%,但均高于体重预期值,具有典型北方小型鸟类的特征(Zheng et al.,2013)。
| 图表编号 | XD00207922900 严禁用于非法目的 |
|---|---|
| 绘制时间 | 2021.01.01 |
| 作者 | 王虹铭、王咏、章熹妮、郑桑榆、高璐瑶、郑蔚虹 |
| 绘制单位 | 温州大学生命与环境科学学院、温州大学生命与环境科学学院、温州大学生命与环境科学学院、温州大学生命与环境科学学院、温州大学生命与环境科学学院、温州大学生命与环境科学学院 |
| 更多格式 | 高清、无水印(增值服务) |


{kind=link}