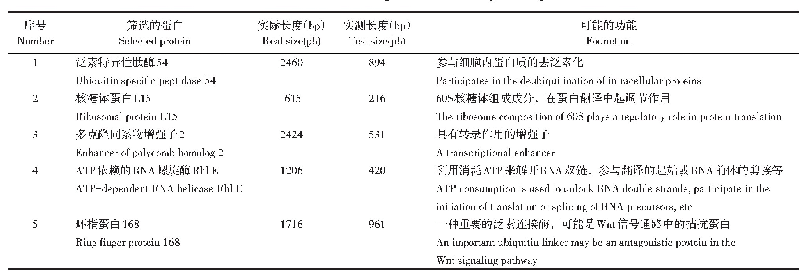
《表2 MED25互作的转录因子》
 提示:宽带有限、当前游客访问压缩模式
提示:宽带有限、当前游客访问压缩模式
MED25不仅参与调控植物的各种器官的发育以及干旱等非生物胁迫,还参与了抗病防御过程。在MED25基因的突变阻断了JA信号途径,会导致其叶片对寄生真菌甘蓝链格孢(A.brassicicola)和灰霉病(B.cinerea)侵染的敏感性提高,但是通过阻止JA信号途径,显示了根对感染半活体真菌尖孢镰刀菌(F.oxysporum)的抗性(Kidd et al.,2009)。事实上,M-ED25突变体在JA途径中,包括PDF1.2、VSP2和MYC2的几个重要基因的诱导表达水平均低于野生型。实验结果显示,MED25作为正调控因子存在于依赖JA信号的防御途径中。根据已有的研究,显示了与MED25互作的转录因子关系(表2),通过对其中的转录因子MYC2和ERF1系统研究后,对MED25是如何在抗病防御过程中发挥作用的机制做了更加清晰的阐述(Ou et al.,2011;覶evik et al.,2012;Chen et al.,2012)。除了MED25,MED8也被报道参与了依赖于JA的防御途径。MED8的突变一方面导致了对甘蓝链格孢(A.brassicicola)的敏感性的增加;另一方面也增加对尖孢镰刀菌(F.oxysporum)的抗性(Kidd et al.,2009)。作为调节植物防御的HUB1因子可与MED21互作,参与植物的防御过程(Dhawan et al.,2009)。在组蛋白H2B单泛素化过程中,HUB1作为其中起重要作用的环状E3连接酶,并在植物防御过程中增强了对甘蓝链格孢(A.brassicicola)和灰霉病(B.cinerea)的抗性(Dhawan et al.,2009)。当拟南芥被白粉菌(E.cichoracearum)侵染或激素诱导后,会导致MED21的积累(Zimmermann et al.,2004;Dhawan et al.,2009)。HUB1的突变体表现出对甘蓝链格孢(A.brassicicola)和灰霉病(B.cinerea)的敏感性。将MED21 RNAi干扰后也表现于hub1突变体相似的性状,而过表达后却与野生型表型上并没有区别(Dhawan et al.,2009)。MED19a是一个调控抵抗霜霉病病原体(Hpa)的正调控因子。med19突变体显示出增加了对Hpa的敏感性。MED19突变后,影响了生物体内的多种防御基因的转录本,在JA/ET信号得以增强,但在SA引起的免疫应答(SAT)方面有所减少(Caillaud et al.,2013)。在拟南芥中NBR4作为M-ED15的同源基因(Canet et al.,2012)。当对med15/nrb4突变体使用SA激素处理后,根据其响应结果显示,MED15与NPR1在功能上表现出高度相似,NPR1是参与植物防御过程中包括SAR等免疫反应的主要调节因子(Canet et al.,2012)。med15/nrb4和npr1的相似性在增强了对疾病敏感性、生物介导的SAR反应(SAR)、效应物触发的免疫反应和非宿主病原体反应。此外,过表达MED15的转基因植株在生长和防御反应方面增强了SA反应。此外,MED15还未被证明在JA信号途径中发挥作用(Canet et al.,2012)。
| 图表编号 | XD0019103900 严禁用于非法目的 |
|---|---|
| 绘制时间 | 2018.12.14 |
| 作者 | 罗骏、巫燕飞、盛锋、杜雪竹 |
| 绘制单位 | 湖北大学生命科学学院、湖北大学生命科学学院、湖北大学生命科学学院、湖北大学生命科学学院 |
| 更多格式 | 高清、无水印(增值服务) |



{kind=link}