《表1 不同元素吸收速度一种变化模型》
 提示:宽带有限、当前游客访问压缩模式
提示:宽带有限、当前游客访问压缩模式
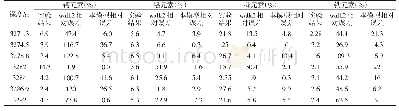
以上述养分吸收模型可产生如下推论。(1)在饱和期,理论上是不存在杂种优势的;或者说,在植株生长很早时期和环境养分供给极端充分时,杂种优势平均值较小或趋于零,变化也较小。可能这个时期很短,以致不易被分辨出来。(2)如果生长时间足够长或各种养分已进入相对胁迫期,植株体养分吸收与环境供给相容性好易获得较大的质量(鲜质量或干质量,下文同),相容性与质量呈正向关系。相容性好指各种养分有较大的饱和吸收速度、和环境供给有较好的协调性即各种养分进入临界点时间相近以及各种养分有较大的(临界)环境消耗率等。由于以相同的外部环境作共同参照,双亲质量越大,则遗传相似性越好;双亲质量越小,由于存在各类不相容性,双亲总体上有较大的遗传差异,但不同组合间可有一定变化。环境相容性好的双亲从通过临界点到其后延续时间里,各种营养元素吸收速度的衰减(相对于饱和吸收速度衰减,下文同)和杂种一代相近,衰减期也和杂种一代较同步,因此不易获得较大的杂种优势;环境相容性差且遗传差异较大时,可导致双亲间各养分平均衰减幅度更大,平均衰减时间更早,易形成较大的杂种优势。因此在大多数情况下,双亲质量之和与杂种优势易呈显著的负相关。(3)植株体质量的变化与饱和养分吸收速度及其衰减均有关。如果前期一直处于饱和期,在临界期附近;或植株生长很早时期和养分供给相对充足条件下,植株质量主要与各种养分尤其是大量元素饱和吸收速度有关,双亲质量之差可一定程度上反映遗传差异;其较大时,双亲之一或双亲养分吸收速度在后续时间内有较大的衰减,衰减时间也较早;因此,此时刻双亲质量之差与邻近较短时段内植株体质量增加量杂种优势可呈正相关。(4)由于植株体质量取样时间、试验环境、遗传背景的巧合,质量较小的亲本或因饱和期养分吸收速度慢未通过或较迟通过临界点,或因饱和期养分吸收速度慢通过临界点后衰减也慢,也可能它们遗传相似性好;质量较大的亲本则相反。但通过临界点之后延续时间均不长,饱和期养分吸收速度主要决定了植株体质量。这时双亲质量之和与杂种优势可呈弱正相关或正相关。但不排除一些不同试验条件下可产生相似的遗传过程。下文通过文献数据的计算结果来证明以上推论。马谦[13]的试验数据表明,大田条件下水稻播种后第20天的幼苗干质量杂种优势为-5.8±19.4(%),而第30天为23.41±19.1(%),第20天到第30天幼苗干质量增加量杂种优势与第20天双亲干质量之差绝对值呈正相关,相关系数为0.379 9(P<0.05),与第30天干质量双亲之和呈负相关,相关系数为-0.527 2(P<0.005)。另外,第20天幼苗干质量杂种优势与同时刻双亲干质量之和相关系数为-0.044 5,第30天幼苗干质量的杂种优势与此时双亲之和呈弱负相关,相关系数为-0.323 7(P<0.1)。以上结果表明,第20天时几乎所有亲本仍处于饱和期或在临界点附近。第30天时,所有亲本均已通过临界期,但其后延续时间均较短或各种养分环境胁迫仍不严重。马谦[13]还有一个黑暗条件之下水稻种子发芽4天后幼芽干质量数据,幼芽干质量的杂种优势与双亲之和呈负相关,相关系数为-0.677 8(P<0.05),杂种一代幼芽干质量和中亲值之差(离亲值,下文同)与中亲值相关系数为-0.603 4(P<0.1),由于发芽时种子贮藏的干物质是唯一能量来源,说明第4天时,各种营养元素已存在明显的胁迫。余丽霞[14]棉花杂交种愈伤组织培养试验数据计算表明,愈伤组织质量杂种优势与同时刻双亲之和的相关系数在第35天、第65天、第80天分别为-0.597 4(P<0.002)、-0.675 0(P<0.001)、及-0.696 5(P<0.001);杂种一代愈伤组织重量离亲值与同时刻中亲值相关系数在第35天、第65天及第80天分别-0.601 0(P<0.002)、-0.670 0(P<0.001)、-0.700 6(P<0.001);第35天到第65天愈伤组织质量增加量杂种优势与第35天、第65天、第80天双亲质量之和相关系数分别为-0.508 2(P<0.01)、-0.714 1(P<0.001)、-0.708 9(P<0.001),说明在第35天时,各种养分早已进入协迫阶段。第65天到第80天愈伤组织质量增加量杂种优势与第35天、第65天、第80天双亲质量之和相关系数分别为-0.544 2(P<0.005)、-0.533 1(P<0.01)、-0.590 3(P<0.002),可能生长后期外界养分已较严重亏缺,杂种一代对各种养分吸收速度也衰减严重,杂种优势变得较不活跃,双亲遗传差异大小与愈伤组织增加量杂种优势相关性减弱。王章奎[15]营养液栽培小麦试验表明,拔节期单株根系干质量杂种优势与双亲之和相关系数为-0.750 5(P<0.001),杂种一代单株根系干重离亲值与中亲值相关系数为-0.704 9(P<0.001),说明这时营养液各种养分的供给已进入较重的胁迫阶段。温玲[16]甜瓜杂交种试验表明,4叶一心期单株根系干质量杂种优势与双亲之和相关系数为0.743 5(P<0.05),这与上文推论第(4)点有关。同时也表明取样时各种养分的环境供给可能仍比较充裕。蔡丽艳等[17]苜蓿杂交种数据也可能属于同样性质,苜蓿干草单株质量的杂种优势与双亲之和相关系数为0.388 3(P<0.05)。大田作物经过一个生长季节,土壤养分已处于相对亏缺状态,这时作物产量杂种优势与双亲之和可为显著或极显著负相关。林欣立等[18]萝卜杂交试验表明,单根质量杂种优势与双亲之和相关系数为-0.583 8(P<0.05)。潘春桂[19]计算结果表明,皮棉产量杂种优势与双亲之和相关系数为-0.696 6(P<0.001)。徐静斐和汪路应[20]的水稻试验表明,水稻产量的杂种优势与双亲之和相关系数为-0.523 4(P<0.002)。司洪华[4]玉米杂交试验表明,玉米产量杂种优势与双亲之和相关系数为-0.840 3(P<0.001)。吴利民等[21]小麦杂种试验,小麦产量杂种优势与双亲之和相关系数为-0.483 3(P<0.05)。梁耀平等[22]西瓜杂交试验表明,西瓜产量杂种优势与双亲之和相关系数为-0.715 7(P<0.001)。阎冬生[23]大豆杂交试验表明,大豆产量杂种优势与双亲之和相关系数为-0.848 5(P<0.001)。桑世飞等[24]的油菜杂交试验表明,油菜产量杂种优势与产量双亲之和相关系数为-0.523 4(P<0.001)。由于杂种优势与双亲之和相关性受到自相关的影响,进一步计算出杂种一代产量离亲值与中亲值的相关性,绝大多数仍呈显著或极显著负相关,只有林欣立、司洪华的试验中两者呈弱负相关,这里未列出相关系数。另外,上文已经提及,双亲之和越小时,双亲遗传差异在不同组合间可变化较大。因此,从线性相关的计算原理可知,由于亲本选配的随意性,有些情况下去除双亲之和最小的一些组合,可显著提高杂种一代离亲值或杂种优势与中亲值相关性。当然与以下原因有关。如果杂交试验中存在很多如下类型组合,因各种养分的吸收速度或(临界)环境消耗率较小导致双亲质量(之和)越小,杂种一代质量由于各种养分进入临界点同步性改善而获得的离亲值反向同步增大受到限制,因此离亲值与中亲值或双亲之和的反相关性减弱或不存在。这在公开发表的文献中十分常见。例如桑世飞等[24]油菜杂交试验原有组合46个,去除产量双亲之和最小的6个组合,杂种一代产量离亲值与中亲值相关系数由原-0.454 8(P<0.002)变为-0.564 3(P<0.005)。王建军等[25]油菜杂交试验原有杂交组合36个,去除籽粒产量双亲之和最小的2个组合,籽粒产量离亲值与双亲之和相关系数由原-0.326 3(P<0.1)增加为-0.474 3(P<0.005)。唐梅[26]水稻杂交试验重庆试点原有组合35个,现去除单株粒质量双亲之和最小的8个组合,离亲值与双亲之和的相关系数由原-0.183 0增加为-0.539 4(P<0.005)。由于植株体主要由大量元素构成,但它对大量元素和少(微)量元素的获取是相互协同和相互制约的关系,因试验环境、遗传背景、目标性状及取样时间不同,杂种优势与亲本生物体质量的相关性存在可解释的变化,但在生长后期或环境养分相对亏缺时,负相关性几乎是必然的,有兴趣的读者可以去验证。
| 图表编号 | XD00172855100 严禁用于非法目的 |
|---|---|
| 绘制时间 | 2020.05.01 |
| 作者 | 潘春桂 |
| 绘制单位 | 安徽省东至县种子管理站 |
| 更多格式 | 高清、无水印(增值服务) |
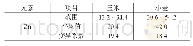



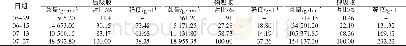
{kind=link}