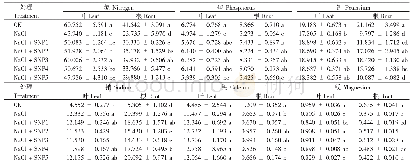
《表1 MCs对水稻幼苗矿质营养含量的影响》
 提示:宽带有限、当前游客访问压缩模式
提示:宽带有限、当前游客访问压缩模式
注:表中数据为平均值±标准差,不同大、小写英文字母代表恢复期、胁迫期处理组间显著性差异(P<0.05)。
矿质营养元素参与细胞的组成、酶的活化和调控生理过程[22]。如表1所示,胁迫7 d后,1μg·L-1MCs组水稻幼苗中矿质营养(K+、Na+、Ca2+、Mg2+、Fe2+、Zn2+、Cu2+、PO43-、NO3-和NH4+)含量无显著变化。10μg·L-1MCs组水稻幼苗中Mg2+、Fe2+、Zn2+、NO3-和NH4+含量增加,PO43-含量降低。推测原因如下:10μg·L-1MCs增强了矿质元素跨膜运输的驱动;植物中矿质元素的吸收与其转运通道及蛋白有关,因此10μg·L-1MCs处理还可能通过影响Mg、Fe、Zn和N转运蛋白家族的表达来增强植物对MCs胁迫的适应性[23-24]。Mg2+、Fe2+和Zn2+是叶绿素生物合成和光合作用的必需元素[25],10μg·L-1MCs胁迫下水稻中Mg2+、Fe2+和Zn2+含量的增加利于维持地上部光合作用以适应MCs胁迫。同时N元素的增加利于提高酶活性调节激素含量,如生长素(IAA)和玉米素(ZT),从而有助于调节水稻的生长[26]。由于植物中Fe、Zn与P存在竞争作用和拮抗作用[27-28],该处理水稻中Fe2+、Zn2+含量的增加可能造成PO43-含量的降低。这些营养元素含量的变化是水稻幼苗对10μg·L-1MCs胁迫的适应性机制之一。100μg·L-1MCs组水稻幼苗中K+、Na+、Mg2+、Zn2+、Cu2+、PO43-和NO3-的含量降低,Ca2+、Fe2+和NH4+含量增加,造成营养元素失衡。Freitas等[29]用100μg·L-1MC-LR处理生菜,10 d后发现叶中矿质元素(Mg、K、P、Mn、Fe、Zn、Cu和Mo)含量均降低。这表明MCs处理下植物中营养物质含量的变化并不一致,可能与植物物种、发育阶段、毒素性质和暴露时间等有关。1000μg·L-1MCs组水稻幼苗中除Ca2+含量增加外,其他元素含量均降低,且降幅大于100μg·L-1MCs组。Ca作为植物信号传导的第二信使,其含量的增加有助于与钙结合蛋白作用,发挥Ca2+的传感器功能,结合基因启动子中的顺式元件来调节植物对逆境的应激反应[30]。结合本项目组前期的报道[8-9],高浓度MCs(100μg·L-1和1000μg·L-1)处理下矿质营养元素的失衡可能与水稻幼苗体内MCs的高积累有关,是造成水稻幼苗光合作用和生物量积累受抑的又一重要原因。
| 图表编号 | XD00109566600 严禁用于非法目的 |
|---|---|
| 绘制时间 | 2019.11.20 |
| 作者 | 刘洪月、申泽辉、梁婵娟 |
| 绘制单位 | 江苏省厌氧生物技术重点实验室江南大学环境与土木工程学院、江苏省水处理技术与材料协同创新中心、江苏省厌氧生物技术重点实验室江南大学环境与土木工程学院、江苏省厌氧生物技术重点实验室江南大学环境与土木工程学院、江苏省水处理技术与材料协同创新中心 |
| 更多格式 | 高清、无水印(增值服务) |




{kind=link}