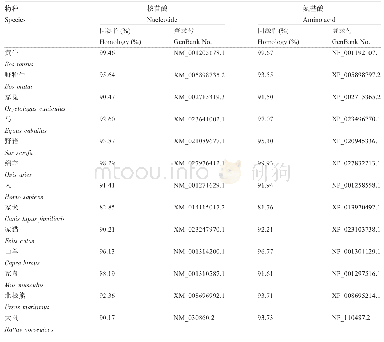
《表1.确定核苷酸序列的tdh基因[9]》
![《表1.确定核苷酸序列的tdh基因[9]》](http://bookimg.mtoou.info/tubiao/gif/WSXB202008004_00500.gif) 提示:宽带有限、当前游客访问压缩模式
提示:宽带有限、当前游客访问压缩模式
本系列图表出处文件名:随高清版一同展现
《副溶血性弧菌耐热性直接溶血素(TDH)的研究进展》
除了在副溶血性弧菌中存在tdh基因外,在其他弧菌属中也发现了tdh基因[6–8]。用tdh基因探针检测发现,所有的霍利斯弧菌(V.hollisae)均含有tdh基因(命名为Vh-tdh);从临床上分离的部分非O1型霍乱弧菌(V.cholerae non-O1)和拟态弧菌(Vibrio mimicus)也携带tdh基因(分别命名为NAG-tdh和Vm-tdh)。通过克隆tdh基因并测序后发现,这3种弧菌的tdh基因和副溶血性弧菌的tdh基因有着非常高的同源性,与tdh2(副溶血性弧菌tdh基因中的一种)的核苷酸序列相似度分别为93.3%、98.6%、97.0%[9](表1)。此外,非O1型霍乱弧菌所携带的NAG-tdh和副溶血性弧菌AQ3776所携带的tdh4(副溶血性弧菌tdh基因中的一种)都位于质粒上,两者的tdh基因核苷酸序列相似度为100%[6],并且在这两种菌株中,携带tdh基因的质粒整体上也有高度同源性。这些结果证明,质粒可介导tdh基因在副溶血性弧菌和霍乱弧菌之间进行转移传播[6]。但是,临床分离的副溶血性弧菌基本上都携带tdh基因,而临床分离的非O1型霍乱弧菌却极少携带tdh基因,所以更倾向于推测tdh基因最初是通过副溶血性弧菌由质粒作为载体转移到非O1型霍乱弧菌当中的。一般情况下,副溶血性弧菌tdh基因位于其2号染色体上的致病岛区域[10](图1),致病岛的获取与水平基因转移或噬菌体有关[11]。因此tdh基因可能通过质粒与染色体在弧菌属之间进行转移传播。在各弧菌属中,tdh基因的侧翼是类插入序列(insertion sequence in vibrios,ISVs)或源自ISVs基因重排的相关序列[12]。ISVs的核苷酸序列与插入序列(insertion sequence,IS)家族中的IS903相似,具有与IS903高度同源的18 bp末端反向重复序列。IS903编码的转座酶具有转座活性,但ISVs编码的转座酶却没有活性。因此,ISVs无法促使tdh基因在弧菌属中进行转移,tdh基因在副溶血性弧菌的染色体上是稳定存在的[12]。有人据此提出如下关于副溶血性弧菌tdh基因起源的假说[9,12]:ISVs序列的G+C含量(43.7%–46.6%)接近弧菌属基因组的平均G+C含量(46.0%–49.0%),但tdh基因的G+C含量(31.5%–33.7%)却明显比前两者要低。ISVs在过去的某个时期可能具有转座活性,通过转座将tdh基因在不同菌种之间进行转移,质粒可能是不同菌种之间tdh基因转移的载体。通过这种机制,tdh基因首先从带有tdh基因的非弧菌菌株转移到弧菌中,然后在不同弧菌之间进行转移。经过了长期的进化,ISVs序列发生了很大的变异,失去了转座活性。另外一个可能的机制是转导,但目前尚没有证据表明噬菌体参与了tdh基因的转移。大多数ISV转座酶基因序列是由于碱基的改变和缺失而发生了改变,在拟态弧菌中发现的一个ISV-M5.2则包含可能编码活性转座酶的开放阅读框(open reading frame,ORF),但未能检测到该转座酶的活性。这些结果表明,引入到弧菌中的tdh基因可能已经在其染色体上固定,而tdh基因两侧对活性的ISVs缺乏选择性压力,这使得这些ISVs容易多样化和缺失,同时tdh基因编码序列变得不易转移与传播。
| 图表编号 | XD00156701100 严禁用于非法目的 |
|---|---|
| 绘制时间 | 2020.08.04 |
| 作者 | 檀利军、王敬敬、石千黛、刘海泉、赵勇 |
| 绘制单位 | 上海海洋大学食品学院、上海海洋大学食品学院、上海水产品加工贮藏工程技术研究中心、农业农村部水产品贮藏保鲜质量安全风险评估实验室(上海)、上海海洋大学食品学院、上海海洋大学食品学院、上海水产品加工贮藏工程技术研究中心、农业农村部水产品贮藏保鲜质量安全风险评估实验室(上海)、上海海洋大学食品学院、上海水产品加工贮藏工程技术研究中心、农业农村部水产品贮藏保鲜质量安全风险评估实验室(上海) |
| 更多格式 | 高清、无水印(增值服务) |



{kind=link}